
Lo scopo del regno vegetale è di nutrire animali e uomini, consolidare il terreno, accrescere la bellezza e mantenere l'equilibrio nell'atmosfera. Mi venne detto che le piante e gli alberi cantano silenziosamente per noi umani e che tutto ciò che chiedono in cambio è di cantare per loro. (Marlo Morgan, "...E venne chiamata Due Cuori")
Post scritto per suggerimento di Mirco, grazie Mirco! Le piante sono organismi viventi pluricellulari, molto importanti per l’ equilibrio degli ecosistemi terrestri e per la vita di tutti gli altri organismi che popolano il nostro pianeta e sono organismi AUTOTROFI.
Le piante sono organismi viventi pluricellulari, molto importanti per l’ equilibrio degli ecosistemi terrestri e per la vita di tutti gli altri organismi che popolano il nostro pianeta e sono organismi AUTOTROFI. AUTOTROFIA vuol dire che ricavano l’energia necessaria per le diverse attività, da sé, attraverso la Fotosintesi Clorofilliana, mentre, l’uomo e gli altri animali sono organismi ETEROTROFI, cioè, hanno bisogno di alimentarsi per ricavare dal cibo, tutta l’energia di cui hanno bisogno. Inoltre, le piante sono considerati organismi PRODUTTORI, nell’ambito dell’ecosistema, in quanto producono l’Ossigeno necessario alla respirazione degli animali, compreso l’uomo e fornisce energia i CONSUMATORI PRIMARI che si nutrono di sole piante e agli organismi ONNIVORI.

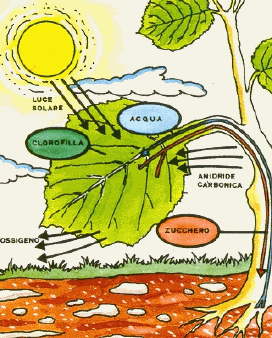
La fotosintesi è il processo biochimico che consente di trasformare l'energia solare, energia termica, in energia chimica. Viene effettuata dalle piante, che contengono la clorofilla, per produrre glucosio, dall'acqua contenuta nel terreno e dall'anidride carbonica presente nell'atmosfera. La clorofilla è un particolare pigmento verde fotosensibile, contenuto all'interno di particolari organelli della cellula vegetale, i cloroplasti, in grado di assorbire la luce solare.
 |
| Cloroplasto |
Anche se tutte le parti verdi di una pianta possono svolgere la fotosintesi, la maggior parte dei cloroplasti si trova nelle foglie: il colore verde è infatti dovuto ai pigmenti di clorofilla, che catturano l’energia luminosa. I cloroplasti sono concentrati nelle cellule del mesofillo, il tessuto verde presente all’interno della foglia. L'anidride carbonica può entrare e l'ossigeno può uscire dalla foglia grazie a piccolissimi pori detti stomi. Le membrane del cloroplasto, una esterna ed una interna separate da uno spazio, formano la rete strutturale in cui avvengono molte reazioni fotosintetiche. La membrana interna dei cloroplasti racchiude un secondo compartimento contenente un liquido denso, lo stroma, in cui vengono sintetizzati gli zuccheri a partire dall’anidride carbonica. Nello stroma sono immersi i tilacoidi, sacchetti discoidali provvisti di membrane, disposti in pile dette grani.
Prima di andare avanti nella fotosintesi, vediamo brevemente come è fatta una foglia dall'interno, dal momento che è il "laboratorio chimico" dove si svolge la fotosintesi.

Esaminando una sezione trasversale di una foglia vediamo che vi sono, sia verso l’alto che verso il basso, uno o due strati di cellule epidermiche, rispettivamente l’epidermide superiore e l’epidermide inferiore, rivestiti da una cuticola molto ispessita nell’epidermide superiore. Compreso fa i due esiste un tessuto parenchimatico, il mesofillo, formato da due zone: la zona più in alto consta di cellule allungate in direzione perpendicolare alla superficie della pagina superiore con molti cloroplasti costituisce il cosiddetto strato a palizzata,mentre la zona inferiore è formata da cellule allungate, con un numero minore di cloroplasti, disposte irregolarmente e in modo da lasciare fra di esse grandi spazi o lacune, questa zona prende il nome di strato lacunoso.
Le sostanze colorate, o pigmenti, che rendono possibile la fotosintesi sono contenute nello spessore della membrana dei tilacoidi. Tali pigmenti sono costituiti da clorofilla, in prevalenza verdi, da carotenoidi che possono essere gialli, rossi, porpora, da Xantofille prevalentemente gialle; da ficobiline blu e rosse. Nella membrana dei tilacoidi, gruppi di questi pigmenti formano, assieme a una catena fotosintetica, ossia a una sequenza di molecole, dei sistemi, trasportatrici di elettroni, altamente organizzati denominati fotosistemi. Esistono due tipi di fotosistemi chiamati rispettivamente fotosistema I e fotosistema II. Ogni fotosistema consiste di:
- Un insieme di circa 300 molecole di clorofilla e altri pigmenti, chiamati pigmenti antenna, perché hanno il compito di assorbire le radiazioni luminose e di convogliarne l’energia verso un centro di reazione associato a una proteina
- Una catena fotosintetica, adiacente al suddetto insieme (della catena fotosintetica fanno parte molecole trasportatrici, come i citocromi, simili a quelli che fanno parte della catena respiratoria)
Nel centro di reazione di ogni fotosistema si trova una particolare molecola di clorofilla, sostanza chiave della fotosintesi, in corrispondenza della quale l’energia della luce viene trasformata in energia chimica. I centri di reazione dei due tipi di fotosistemi I e II , assorbono differenti lunghezze d’onda. La clorofilla del centro di reazione del fotosistema I viene denominata P700 per indicare che assorbe onde luminose con lunghezza d’onda di 700 nm, mentre la clorofilla del centro di reazione del fotosistema II viene denominata P680 perché assorbe onde luminose con lunghezza d’onda di 680 nm . I due fotosistemi sono strettamente associati fra loro. L’insieme delle numerose reazioni, catalizzate da enzimi, che costituiscono la fotosintesi può essere suddiviso in due fasi, denominate rispettivamente fase luminosa e fase oscura. Nelle reazioni della fase luminosa, i due fotosistemi della membrana dei tilacoidi catturano l’energia luminosa e convertono parte di essa in energia chimica di molecole trasportatrici di energia, l’ATP e il NADPH. Quest’ultimo differisce dal NADH per un gruppo fosfato in più legato a uno dei gruppi –OH del ribosio. Nelle reazioni della fase oscura, enzimi presenti nello stroma utilizzano l’energia dell’ATP e del NADPH, prodotti nella fase luminosa, per realizzare la sintesi del glucosio.
Le fasi della fotosintesi
La fase luminosa e analogie con la fosforilazione ossidativa
Nella fase luminosa vengono prodotti ATP e NADPH. Per comprendere gli eventi della fase luminosa può essere utile individuare analogie e differenze con la fosforilazione ossidativa:
Entrambi i processi avvengono nello spessore di membrane biologiche: la fosforilazione ossidativa nella membrana interna dei mitocondri; le reazioni della fase luminosa nella membrana dei tilacoidi e dei cloroplasti
Nel corso della fosforilazione ossidativa, gli elettroni e gli ioni H+ provenienti dal metabolismo ossidativo e trasportati dal NADH, vengono ceduti a una catena respiratoria: attraverso questa, gli elettroni vengono trasferiti all’ossigeno che si riduce a ione ossigeno O2-, che si combina con due ioni H+ del mezzo ambiente, formando H2O secondo la reazione:
½ O2 + 2 e- + 2 H+ → H2O
Contemporaneamente, gli ioni H+ vengono pompati dalla matrice al compartimento esterno del mitocondrio: si forma così una differenza di concentrazione di ioni H+ che costituisce una fonte di energia potenziale, utilizzata per fosforilare l’ADP in ATP. Inoltre al momento in cui alimenta la catena respiratoria il NADH si ossida a NAD+. Nel corso delle reazioni della fase luminosa avviene praticamente l’inverso: due elettroni e due ioni H+ vengono allontanati da una molecola di H2O per cui l’ossigeno viene ossidato a ossigeno elementare, secondo la reazione:
H2O→ 2 e- + 2 H+ + ½ O2
Questa scomposizione dell’acqua, che avviene grazie all’energia della luce, viene denominata fotolisi. L’ossigeno che si forma nella fotolisi si libera come prezioso sottoprodotto del processo foto sintetico. Gli ioni H+ che si formano nella fotolisi restano in soluzione nel mezzo ambiente, mentre gli elettroni vengono avviati, attraverso i due foto sistemi e le relative catene foto sintetiche, verso una molecola di NADP+, che viene ridotta, anche con l’intervento di H+ a NADPH. Contemporaneamente, lungo una delle catene foto sintetiche, gli ioni H+ vengono pompati dallo stroma al lume interno dei tilacoidi: si forma così una differenza di concentrazione di ioni H+ che costituisce una differenza di energia potenziale, utilizzata per trasformare l’ADP in ATP. Questo processo avviene ad opera dell’enzima ATP fosforilasi, che fa parte di un sistema canale-enzima, del tutto analogo a quello osservabile nei mitocondri, e che corrisponde alle escrescenze presenti sulla superficie esterna della membrana dei tilacoidi. Nel caso dei cloroplasti, dato che la produzione di ATP avviene grazie all’energia della luce, il processo di formazione di questa sostanza viene denominato fosforilazione
Il NAD+ prodotto dalla fosforilazione ossidativa viene riutilizzato per la demolizione del glucosio; viceversa il NADPH prodotto nelle reazioni della fase luminosa viene utilizzato per la sintesi del glucosio.
Nella catena respiratoria della fosforilazione ossidativa, si ha un processo interamente “ in discesa”: le molecole trasportatrici di elettroni sono disposte, infatti, secondo valori crescenti del potenziale di riduzione e gli elettroni vengono facilmente trasferiti da una molecola all’altra della catena, fino all’accettore finale l’ossigeno. Nella sequenza costituita dai due foto sistemi e dalle due annesse catene fotosintetichie, si ha una situazione meno semplice, come mostra lo schema:

Per la sua forma , è tradizionalmente denominato schema Z. Lo schema Z è essenzialmente un grafico dell’andamento dell’energia libera lungo i due fotosistemi e le relative catene fotosintetiche. Osservando la figura si può notare che nel caso dei cloroplasti, gli elettroni devono trasferirsi complessivamente “contro corrente”. Infatti anche se le due catene fotosintetiche della sequenza sono in discesa, il percorso completo degli elettroni prevede uno spostamento dall’acqua, al NADP+. Questo comporta per gli elettroni un salto energetico che può essere compiuto solo con un sufficiente apporto di energia libera dall’ambiente. L’unica fonte praticamente illimitata di energia libera è il Sole: i cloroplasti provvedono a catturarne quanta ne occorre mediante i pigmenti antenna dei fotosistemi I e II. Fatte queste premesse, possiamo esaminare più in dettaglio il funzionamento dei due fotosistemi. Può essere utile a tal fine, seguire il percorso degli elettroni, dalla molecola di H2O al NADP+, tenendo conto della disposizione dei vari componenti nella membrana di un tilacoide. Quando un fotone, convogliato dai pigmenti antenna, raggiunge la molecola di clorofilla che costituisce il centro di reazione del fotosistema II, la sua energia si trasforma in energia chimica, in quanto viene assorbita da uno degli elettroni della molecola di clorofilla. L’elettrone così eccitato abbandona la molecola di clorofilla e viene catturato dalla prima molecola trasportatrice dell’adiacente catena fotosintetica. La molecola di clorofilla, privata dell’elettrone, resta in un stato ossidato, avido di elettroni. Mediante un meccanismo enzimatico essa recupera un elettrone da una molecola di acqua presente nel lume interno del tilacoide, dopo di che è pronta a svolgere nuovamente la sua funzione di centro attivo. Man mano che l’elettrone passa da un trasportatore all’altro, lungo la catena, perde l’energia in più fornitagli dal fotone.
Uno dei trasportatori di questa prima catena fotosintetica, oltre a trasferire l’elettrone al trasportatore successivo, pompa uno ione H+ dallo stroma al lume interno del tilacoide. Una volta giunto al termine della prima catena fotosintetica, l’elettrone ormai a un livello molto basso di energia libera, viene trasferito al fotosistema I. Ciò è possibile solo se la molecola di clorofilla nel centro di reazione P700 si trova allo stato ossidato, cioè è stata a sua volta raggiunta da un fotone che l’ha privata di un elettrone, eccitandolo. Pertanto, mentre la molecola di clorofilla nel centro di reazione P680 viene ridotta da un elettrone proveniente da una molecola di acqua, quella del centro di reazione P700 viene ridotta dall’elettrone proveniente dal centro di reazione P680. Il viaggio attraverso la seconda catena fotosintetica dell’elettrone che ha lasciato il foto sistema I ha come unico scopo la riduzione di una molecola di NADP+ a NADPH. In sintesi, la serie di eventi descritti può essere vista come un unico processo in cui gli elettroni fluiscono, per azione dell’energia luminosa dall’acqua al NADP+.
Nella fase oscura che segue l’energia delle molecole di ATP e di NADPH prodotte nella fase luminosa viene utilizzata per sintetizzare il glucosio a partire da biossido di carbonio. La denominazione fase oscura riferita alla serie di reazioni necessarie per il completamento della fotosintesi non sta ad indicare che le reazioni debbano avvenire al buio, ma piuttosto che esse possono avvenire anche in assenza di luce. Infatti questa seconda fase del processo foto sintetico non richiede l’intervento della luce: le molecole di ATP e NADPH prodotte nella fase luminosa forniscono l’energia e i reagenti necessari per ultimare il processo foto sintetico, anche se quest’ultimo è fortemente endoergonico.
La fase oscura della fotosintesi
Le reazioni della fase oscura fanno parte di un ciclo detto ciclo di Calvin che si svolge nello stroma del cloroplasto.
Il ciclo di Calvin è analogo al ciclo di Krebs in quanto, a ogni giro, il composto iniziale viene rigenerato. Il composto iniziale (e anche quello finale) del ciclo di Calvin è costituito da uno zucchero a cinque atomi di carbonio legato a due gruppi fosfato, il ribulosio difosfato (RuDP), che fu scoperto nelle alghe e nelle foglie di spinacio.
Il ciclo comincia quando l'anidride carbonica si lega al RuDP. Per identificare il prodotto di questa reazione, Melvin Calvin e i suoi collaboratori fecero crescere per un breve periodo di tempo alcune alghe fotosintetiche in presenza di molecole di anidride carbonica i cui atomi di carbonio erano stati resi radioattivi. Il primo prodotto in cui trovarono gli atomi di carbonio marcati fu l'acido fosfoglicerico (PGA).
Ciò indicava che, quando l'anidride carbonica si lega, al RuDP, la molecola si scinde immediatamente per formare due molecole di PGA.
Questa reazione è catalizzata da un enzima specifico, la RuDP-carbossilasi oggi nota come rubisco. Essa è ritenuta la proteina più abbondante sulla, Terra. Ogni molecola di PGA prodotta in questa reazione iniziale, contiene tre atomi di carbonio; per questo motivo, il ciclo di Calvin è conosciuto anche come via del C3.
Come nel ciclo di Krebs, ogni passaggio del ciclo di Calvin è catalizzato da un enzima specifico. A ogni giro completo una molecola di anidride carbonica entra nel ciclo, viene ridotta e si riforma una molecola di RuDP tre giri del ciclo fanno entrare tre molecole di anidride carbonica, sufficienti per ottenere uno zucchero a tre atomi di carbonio, la gliceraldeide 3-fosfato (G3P).
6 CO2 + 6 H2O → C6H12O6 + 6 O2

Guardando la figura, partendo dalla scritta in alto a sinistra, potete notare che sei molecole di ribulosiodifosfato (RuDP) , un composto a cinque atomi di carbonio, si combinano con sei molecole di anidride carbonica. Ciò produce sei molecole di un composto intermedio instabile che si scinde immediatamente dando origine a dodici molecole di acido fosfoglicerico (PGA), up composto a tre atomi di carbonio.
Nelle successive due tappe queste molecole sono ridotte a dodici molecole di gliceraldeide 3-fosfato. Dieci di queste molecole a tre atomi di carbonio si combinano e si riassemblano per riformare sei molecole di RuDP a cinque atomi di carbonio (ossia, il composto iniziale). Le due molecole «in più» di gliceraldeide 3-fosfato rappresentano il guadagno netto del ciclo di Calvin. La gliceraldeide 3-fosfato è la stessa molecola a tre atomi di carbonio che si forma quando il fruttosio difosfato si scinde nella quarta tappa della glicolisi (vedi capitolo 8). Usando questa molecola come punto di partenza, la cellula può sintetizzare una certa varietà di zuccheri, amminoacidi e acidi grassi.
Una soluzione alla carenza di CO2 : la fotorespirazione
In presenza di una grande quantità di anidride carbonica, il ciclo di Calvin svolge in genere regolarmente. Invece, quando nella foglia la concentrazione di anidride carbonica è bassa, se paragonata a quella dell'ossigeno, l'enzima RuDP-carbossilasi catalizza una reazione tra RuDP e ossigeno invece che tra RuDP e anidride carbonica.
Questa reazione è la prima tappa di un processo conosciuto come fotorespirazione, in cui i carboidrati vengono ossidati in presenza di luce con formazione di acqua e dell' anidride carbonica. In altre parole, la cellula si autoproduce l'anidride carbonica di cui necessita. Tuttavia, in modo diverso dalla respirazione dei mitocondri, la fotorespirazione non è un processo redditizio dal punto di vista energetico, in quanto non produce né ATP né NADH. In alcune piante più del 50% del carbonio fissato nella fotosintesi può essere riossidato ad anidride carbonica durante il processo di fotorespirazione.
Le condizioni che possono innescare la fotorespirazione sono piuttosto comuni. l'anidride carbonica non è sempre disponibile per le cellule fotosintetiche di una pianta. Come abbiamo visto, questo gas entra nella foglia attraverso gli storni, pori specializzati che si aprono e si chiudono in seguito alla pressione dell'acqua e ad altri fattori. Quando una pianta si trova in un ambiente caldo e asciutto deve chiudere i suoi storni per conservare l'acqua; ciò impedisce il rifornimento di anidride carbonica e favorisce invece l'accumulo di ossigeno prodotto dalla fotosintesi. Come conseguenza si hanno concentrazioni basse di anidride carbonica e alte di ossigeno, fattori che inducono la fotorespirazione.
Una seconda soluzione: la via del C4
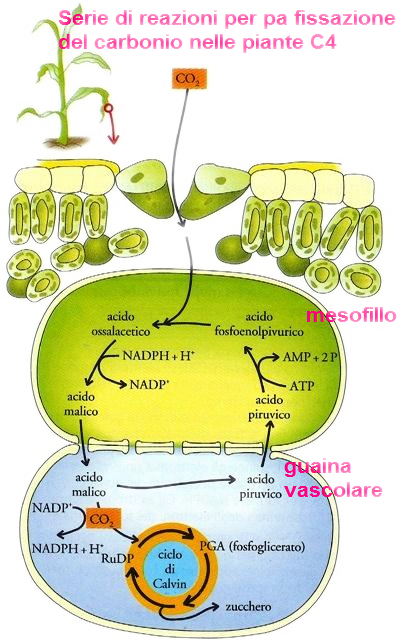
Il problema della carenza di CO2 viene risolto da alcune piante grazie a una via alternativa per catturare questo gas. In queste piante la prima tappa della fissazione del carbonio consiste nel legame tra l'anidride carbonica e un composto chiamato acido fosfoenolpiruvico (PEP), e nella formazione di un composto a quattro atomi di carbonio, l'acido ossalacetico.
Inizialmente, nelle cellule del mesofillo della foglia, l'acido ossalacetico viene trasformato in acido malico
Attraverso i plasmodesmi l'acido malico passa nelle cellule della guaina vascolare, che formano un rivestimento intorno ai fasci vascolari e che, a differenza delle cellule della guaina delle piante C3, contengono i cloroplasti. Qui l'acido malico è demolito in acido piruvico e anidride carbonica, che entra nel ciclo di Calvin l'acido piruvico passa attraverso i plasmodesmi verso le cellule del mesofillo, dove viene ritrasformato in acido fosfoenolpiruvico, pronto per ricevere un' altra molecola di anidride carbonica.
Le piante che utilizzano questa via sono comunemente chiamate piante C4, per distinguerle dalle piante C3 in cui inizialmente il carbonio viene catturato per formare un composto a tre atomi di carbonio, l'acido fosfoglicerico (PGA).
La PEP-carbossilasi, l'enzima che catalizza la sintesi di acido ossalacetico nelle piante C4, ha un' affinità maggiore per l'anidride carbonica rispetto alla RuDP-carbossilasi, l'enzima che catalizza la formazione di PGA. Se gli stomi devono rimanere chiusi gran parte del loro tempo, come accade, per esempio, nei climi caldi e secchi per conservare acqua, la pianta con metabolismo C4 prenderà più anidride carbonica a ogni, per così dire, «respiro», rispetto alla pianta con metabolismo C3. Le piante C4 si sono evolute principalmente nelle zone tropicali e si sono adattate particolarmente bene a un'alta intensità luminosa, alle alte temperature e alla siccità; la canna da zucchero e il granturco sono tra le piante C4 più conosciute.
Una terza soluzione: le piante CAM

Di solito gli stomi delle piante si aprono durante il giorno e si chiudono durante la notte, ma in alcune specie accade esattamente il contrario. Di notte, non solo la temperatura è più bassa, ma l'umidità è in genere più alta; entrambi questi fattori riducono la velocità di traspirazione. Le specie che aprono gli storni soltanto di notte comprendono una certa varietà di piante adattate ai climi caldi e secchi, come, per esempio, i cactus, gli ananas e i membri della famiglia delle crassulacee come, per esempio, il genere Sempervivum che cresce in montagna.
Queste piante assorbono anidride carbonica di notte, trasformandola in acidi a quattro atomi di carbonio; durante il giorno, quando gli stomi si chiudono, l'anidride carbonica viene liberata da questi acidi organici e utilizzata immediatamente per la fotosintesi. Questo processo è detto metabolismo acido delle crassulacee e le piante che lo compiono sono dette piante CAM. Il metabolismo acido è analogo alla via del della fotosintesi, sebbene si sia evoluto probabilmente in maniera indipendente.
Le piante grasse hanno una crescita lenta che è la diretta conseguenza del loro particolare metabolismo. Fino ad ora abbiamo analizzato i metabolismi C3 e C4 che rappresentano delle vie metaboliche “rapide” che portano alla fissazione di CO2 in elementi organici direttamente utilizzabili dalla pianta. Abbiamo avuto modo di dire che la temperatura elevata e la traspirazione rappresentano dei fattori limitanti per molte specie; alcune piante, tra cui quelle grasse, si sono adattate a climi torridi modificando il loro metabolismo. Se nelle piante C4 si assiste ad una separazione spaziale della funzionalità carbossilative nelle CAM si apprezza una separazione temporale nel senso che gli eventi di acquisizione di anidride carbonica e di fissazione di questa molecola risultano temporalmente separati tra la notte ed il giorno. Per evitare di perdere preziosa acqua nel torrido caldo mattutino le piante CAM, nel corso dell'evoluzione, hanno diminuito la loro effettiva superficie fogliare assumendo forme particolari. Inoltre le CAM lasciano gli stomi chiusi di giorno perché, in questo modo, non fuoriesce il prezioso vapore acqueo nelle torride giornate.
L'apertura degli stomi avviene di notte, quando la temperatura è più bassa ed è possibile l'entrata della CO2 attraverso gli stomi senza una eccessiva traspirazione. L'anidride carbonica viene usata per carbossilare il fosfoenolpiruvato, che deriva dalla glicolisi dell'amido, in ossalacetato che viene ridotto a malato proprio come abbiamo visto nelle piante C4. Il malato viene pompato, contro gradiente, all'interno del vacuolo per evitare che la sua presenza possa acidificare il citosol cellulare. Durante tutto la notte, con gli stomi aperti, si riempiono i vacuoli di malato fino alla loro massima capienza.
Con le prime ore del mattino gli stomi iniziano a chiudersi in risposta all'aumento del lavoro della fase luminosa, con conseguente sintesi di ATP e NADPH, ed il malato inizia ad essere liberato dal vacuolo. L'enzima malico processa il malato che, decarbossilando, forma il piruvato. Il piruvato viene processato dall'enzima piruvatofosfato dichinasi per formare il fosfoenolpiruvato che, nella via glicolitica, torna a formare amido.
Gli enzimi sono sensibili alla concentrazione idrogenionica perché un pH citoplasmatico che vira verso l'acido è indice che il malato non è più conservabile nel vacuolo che risulta saturo. Un pH acido, nel citoplasma, inibisce la PEP-carbossilasi che, in questo modo, non carbossila il fosfoenol-piruvato. Viceversa un pH acido attiva l'enzima malico che processa il malato e, durante il giorno, fornisce CO2 che nel cloroplasto viene fissata nel ciclo di Calvin.
FONTI:
Ciclo di Calvin
La fotosintesi: un sistema redox per la vita
Il metabolismo acido delle crassulacee (CAM)
PER APPROFONDIRE:
Fasi della fotosintesi clorofilliana
Fotosintesi
Una luce sull'origine della fotosintesi

Nessun commento:
Posta un commento